Notes For All Chapters Biology Class 11 CBSE
This chapter introduces the internal structure and functional organisation of higher plants. The study of internal structure of plant is called anatomy.
1. Tissues and Tissue Systems
The plants have cells as their basic unit. Compared to animal cells, plant cells have a cell wall consisting of a primary cell wall, secondary cell wall and middle lamella. To make the plant structure, the cells with common function, joint together and form a complex structure, called tissue.
Tissues
A tissue is a group of cells having a common origin and usually perform a common function. A plant body is made up of different kinds of tissues. Generally, the cells of a tissue share the same origin in the embryonic stage. The tissues help in body function by allowing division of labour, e.g., In leaf, various cells commonly perform the function of photosynthesis.
The plant tissues can be divided into two main types I. Meristematic tissues II. Permanent tissues
I. Meristematic Tissues
The growth in plants is mainly restricted to specialised regions of active cell division called meristems (Gk. Memtar—divided). A meristematic tissue is an undifferentiated mass of cells, that is in a continuous state of division or retain their power of division. These tissues divide to form new cells which differentiate to give rise to permanent tissues.
Characteristics of Meristematic Tissue
The characteristics of meristematic tissue are listed below
(i) They are living and contain undifferentiated mass of rapidly dividing cells.
(ii) The shape of cells is spherical, polygonal or rectangular.
(iii) The cells are compactly arranged without intercellular spaces and are interconnected by plasmodesmata.
(iv) Nucleus is large and present either in interphase or in divisional stages.
(v) Cell wall is thin with only a primary wall made up of cellulose. Secondary wall is absent.
Classification of Meristematic Tissue
Meristem can be classified broadly based on three ways, i.e., position in the plant body, functions and origin.
Classification Based on Position
Meristems can be divided into three types, based on their position in the plant body. These are as given below
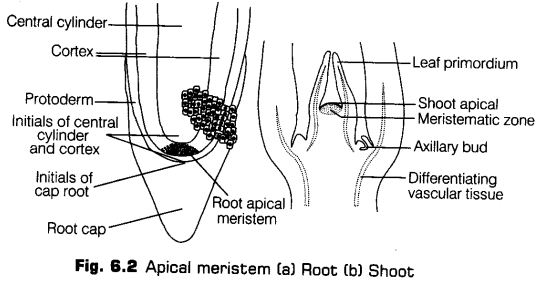
(a) Apical Meristems The meristems which occurs at the tips of root and shoot and produce primary tissues are called apical meristems. The Root Apical Meristem (RAM) occupies the tip of a root while, the Shoot Apical Meristem (SAM) occupies the distinct most region of the stem axis.
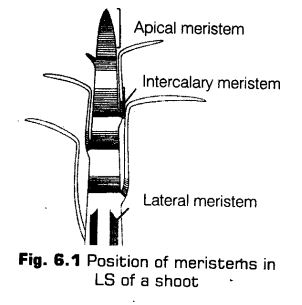
During the formation of leaves and elongation of stem, some cells, left behind from shoot apical meristem, constitute the axillary bud. These buds are present in the axil of leaves and are capable of forming a branch or a flower.
(b) Intercalary Meristems The meristem which occurs between mature tissues is known as intercalary meristem. They occur in grasses and regenerate parts removed by the grazing herbivores.
Both apical and intercalary meristems are primary meristems because they appear early in life of a plant and contribute to the formation of the primary plant body. These meristems are usually responsible for growth in length and present mostly at the base of node (e.g., Mint), base of internode (e.g., Stem of wheat and grasses) or at the base of leaf (e.g., Pinus).
(c) Lateral Meristems The meristems that occurs in the mature regions of roots and shoots of many plants. These meristems produce woody axis and appear later than primary meristem is called the secondary or lateral meristem.
They are cylindrical meristems. Some examples of lateral meristems are fascicular vascular cambium, interfascicular cambium and cork cambium. These are responsible for producing the secondary tissues.
Classification Based on Functions
The meristems are also classified on the basis of their functions as
(a) Protoderm It is the outermost portion of the primary meristem found at the apex of the stem and root. It develops into epidermis.
(b) Procambium It develops into primary vascular tissues. It forms the isolated strands of elongated cells, very near to the central region.
(c) Ground Meristem It develops into the ground tissue. The cells are thin-walled, living and isodiametric. In the later stages of growth, they become differentiated into hypodermis, cortex, endodermis, pericycle, medullary rays and pith.
Classification Based on Origin
The meristems are grouped on the basis of origin as
(a) Primary Meristems These meristems are derived during the early embyonic stages. They divide rapidly and differentiated into primary permanent tissues which make the fundamental structure of the plant body. They are mainly found in the growing apical regions of the root and shoot.
(b) Secondary Meristems These meristems appears in later stage of development in the plant body. They lie lateral in position in both the stem and root. Some primary permanent tissues acquire the power of division and become meristematic.
These tissues dedifferentiate and form secondary meristems. Some examples of secondary meristems are cork cambium and interfascicular cambium. They allow secondary growth in tissues.
II. Permanent Tissues
The meristematic cells gradually differentiate and become mature or permanent. The permanent tissues actually composed of cells in which the growth has stopped.
Permanent tissues can be divided into two types
1. Simple Permanent Tissues
The permanent tissues having all cells similar in structure and function are called simple tissues. These are groups of homogenous cells which perform the same function.
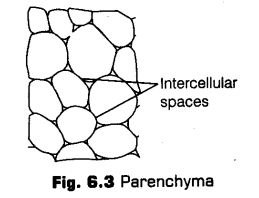
i.Parenchyma
It is a living, simple permanent tissue composed of thin-walled cells. Parenchyma (Para-beside; enchein – to pour) is also called primary tissue or ground tissue. It is present in cortex, pith, palisade, mesophyll and some other parts of flower. It is mostly produced by the ground tissue. The parenchyma terms the major component within the organs.
The characteristic features of parenchyma are as follow
(a) The cells are thin-walled, less spherical and polyhedral in shape, these are generally isodiametric.
(b) The cell wall composed of cellulose, hemicellulose and pectin.
(c) Cells have a large central vacuole, peripheral cytoplasm with a nucleus.
(d) The cells may be either closely packed or have small intercellular spaces.
The parenchyma can be further classified as
* Chlorenchyma specialised for photosynthesis.
* Aerenchyma forms a connected air system throughout the entire plant.
* Storage parenchyma store sugars, protein granules, oil drops, etc.
* Xylem parenchyma helps in the conduction of water.
* Phloem parenchyma help in the translocation of food.
* Stellate parenchyma star-shaped parenchymatous tissue with large air spaces.
Different functions performed by parenchyma are
(a) These helps in storage of food, water and air,
(b) The vital activities like photosynthesis, respiration and conduction are carried out by parenchyma.
(c) It helps in wound healing, grafting, etc., and also provides buoyancy in aquatic plants.
(d) Parenchyma cells associated with xylem and phloem help in conduction of water, and food materials.
(e) These cells can dedifferentiate, acquire the power of division to form secondary meristem which produce secondary tissues.
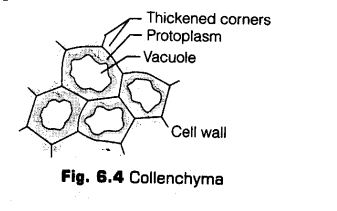
ii. Collenchyma
Collenchyma (Gr. Colla – glue; enchyma – an infusion) is a simple, living mechanical tissue. Its cells composed of more or less elongated cells with thick, primary non-lignified walls. Intercellular spaces are found to be absent.
The characteristic features ofcollenchymatous tissues are listed below
(a) It is present only in the aerial parts of the plant body.
(b) It is found either as a homogeneous layer or in patches.
(c) Collenchyma consists of cells’ which are much thickened at the corners due to a deposition of cellulose, hemicellulose and pectin. id) The cells may be oval, spherical or polygonal and often contain chloroplasts.
(e) These cells assimilate food, when they contain chloroplasts.
Based on pectinisation of the cell wall, there are three types of collenchyma
(a) Angular collenchyma
(b) Lamellar collenchyma
(c) Lacunar collenchyma
Different junctions performed by collenchyma are
(a) It provides mechanical support to the growing parts of the plant, such as young stem and petiole of a leaf.
(b) Collenchyma cells are capable of photosynthesis, as they contain chloroplasts.
iii. Sderenchyma
The sderenchyma (Gr. Sclerous—hard; enchyma—an infusion) consists of long, narrow cells with thick and lignified cells walls having a few or numerous pits.
The characteristic features ofsclerenchymatous cells are
(a) Cells are long or short, narrow, thick-walled and lignified.
(b) They possess hard and extremely thick secondary walls due to uniform deposition of lignin.
(c) These are dead cells and do not perform any metabolic function.
(d) They show different types of lignin depositions and also have pits.
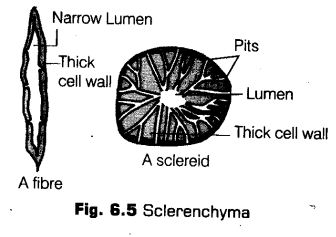
The sclerenchymatous cells may be divided into two types
(a) Sclereids These are short or irregular, spherical, oval or cylindrical sclerenchymatous cells. The walls are very thick, irregular and the lumen is very narrow. The walls show simple pits. These are commonly found in the fruit wall of nuts, pulp of the fruits, like-guava, pear and sapota, seed coats of legumes and leaves of tea.
(b) Sclerenchymatous Fibres These are thick-walled, elongated and pointed cells, generally occurring in groups, in various parts of the plant.
Different functions performed by sderenchyma are
(a) It provides mechanical strength and support.
(b) Surface fibres help in dispersal of seeds.
2. Complex Permanent Tissues
Complex permanent tissues are a group of more than one type of cells having common origin and working together as a unit.
The main complex tissues in vascular plants are xylem and phloem.
Xylem
Xylem (Gr. Xylos — wood) is a complex permanent tissue which conducts water and mineral nutrients upwards from the root to the leaves.
The xylem tissues are composed of four components
a. Tracheids These are elongated, tubular and primitive cells with tapering end walls. They are dead cells and do not contain protoplasts. The inner layers of the cell walls have thickenings which vary in form. The end of the tracheids are tapering, blunt or chisel like. These are constituents of xylem of primitive plants.
These are found in pteridophytes and gymnosperm tracheids may be classified as annular or helical, spiral and scalariform or pitted.
The tracheids conduct water and dissolved mineral elements from roots to leaves. They also provide mechanical support.
b. Vessels These are long, cylindrical, tube-like structures made up of many cells called vessel members, each with lignified walls and a large central cavity.
The vessel cells are also devoid of protoplasm. The vessel members are interconnected through perforations in their common walls. The presence of vessels is a characteristic feature of angiosperms.
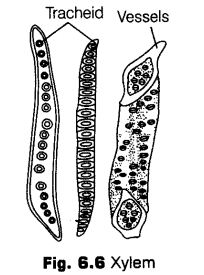
c. Xylem Fibres The sclerenchymatous fibres associated with the xylem are called xylem fibres. These fibres have lignified cell walls. The thickness of the walls varies considerably, but these are usually thicker than the walls of the tracheids in the same wood. These are found in both primary and secondary xylem. The xylem fibres provide mechanical strength.
d. Xylem Parenchyma The parenchyma cells associated with the xylem form xylem parenchyma. These cells form the only living component of the xylem. Xylem parenchyma stores food in the form of starch.
These cells assist direcdy or indirecdy in the conduction of water upward through the vessels and tracheids.
The xylem parenchyma can be sub-divided into two types Primary Xylem The xylem differentiating in the primary plant body is the primary xylem. The primary source of this xylem is the procambium. The primary xylem is of two types, i.e., protoxylem and metaxylem.
The first formed primary xylem elements are called protoxylem.The latter formed primary xylem is called metaxylem.
In stems, the protoxylem lies towards the centre (pith) and the metaxylem lies towards the periphery of the organ. This type of primary xylem is called endarch.
In roots, the protoxylem lies towards periphery and metaxylem lies towards the centre. Such arrangement of primary xylem is called exarch.
Secondary Xylem is composed of tracheary elements, rays, fibres and interspersed axial parenchyma cells. The cell formed toward inside of cambia are called secondary xylem or wood. The primary function of secondary xylem is to provide mechanical support to plants.
ii. Phloem
Phloem (Gk. Phbis—bark) is a food conducting complex permanent tissue. The term ‘phloem’ was coined by Nageli (1958). In angiosperms, it is also called bast. In gymnosperms, albuminous cells and sieve cells are present. The first formed primary phloem consists of narrow sieve tubes called protophloem and the latter formed phloem has bigger sieve tubes called metaphloem.
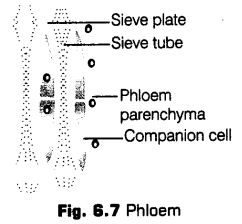
It consists of four types of cellular components,
(a) Sieve Elements The sieve tube elements are long, tube-like structures arranged longitudinally and are associated with the companion cells. Their end walls are perforated in a sieve-like manner to form the sieve plates. A mature sieve element possesses a peripheral cytoplasm and a large vacuole, but lacks a nucleus, Golgi body and most cytosol.
Sieve elements are of following two types
• Sieve cell It is a special kind of cell which posses sieve areas in its lateral walls. There is no specialised plate in it. Sieve cells are usually found in pteridophytes and gymnosperms.
• Sieve tube members In this type, the sieve areas are localised on its end walls. Sieve tube members are placed one above the other forming a continuations tube called sieve tube. The end walls are perforated (sieve pores) like a sieve. These are found in angiosperms.
The uniqueness of the sieve tube is that although without nucleus, it is living and the nucleus of the companion cell controls its functions.
The main function of sieve element is trans¬location of organic solutes. The callose (a plant polysaccharide) is present in the perforations in the sieve plates.
It is soluble and disappears when the solute is dilute so that the solute can pass from one cell to another cell through the pores. Callose reappears and sometimes closes the pores when solute is less dilute, thus stopping the movement.
(b) Companion Cells These are specialised parenchymatous cells, which are closely associated with the sieve tube elements. Usually, a single companion cell is found associated with a sieve tube member.
The cytoplasm of the sieve tube element and companion cells are connected by thin cytoplasmic strands called plasmodesmata, passing through the pit membranes in their walls. Companion cells are absent in the phloem of pteridophytes and gymnosperms. They have albuminous cells.
The companion cells In association with phloem parenchyma play an important role in the maintenance of a pressure gradient in sieve tubes. They form a link between sieve tube cells and other cells and regulate the passage of materials.
(c) Phloem Parenchyma The phloem parenchyma is made up of elongated, tapering cylindrical cells which have dense cytoplasm and nucleus. The cell wall is composed of cellulose and has pits though the plasmodesmatal connections, which exist between the cells.
They store food materials and other substances like resins, latex and mucilage. The phloem parenchyma is absent in most of the monocotyledons.
(d) Phloem Fibres The phloem fibres (bast fibres) are made up of sclerenchymatous cells. These are generally absent in the primary phloem but are found in secondary phloem. The cell wall of phloem fibres is quite thick. At maturity, these fibres lose their protoplasm and become dead. The phloem fibres of jute, flax and hemp have important economic uses.
The Tissue System
The tissues also vary, depending upon their location in the plant body. Their structure and function would also be dependent on location. Thus, on the basis of their structure and location, there are three types of tissue system, i.e., epidermal tissue system, ground or fundamental tissue system and vascular or conducting tissue system.
1. Epidermal Tissue System
The epidermal tissue system forms the outermost covering of the whole plant body. Its various components are epidermal cells, stomata and the epidermal appendages, i.e., trichomes and emergences.
Epidermis The epidermis (Gr. Epi-upon\ derma -skin) is the outermost layer of the primary plant body. The epidermal cells vary in shape and size and are compactly arranged to form a continuous layer. This layer is interrupted by stomata. Sometimes they are separated by intercellular spaces. It is usually single-layered but is also multilayered in the aerial roots of orchids and leaves of Nerium and Ficus elastica.
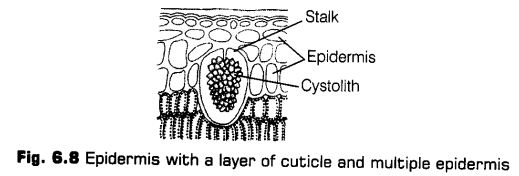
The cells are parenchymatous and living. Each cells has a large central vacuole and a peripheral thin cytoplasm. It is thicker in xerophytic plants. In roots the. outermost layer called epiblema, has tubular, unicellular, projections called root hair. The other substances deposited on cuticle surface may be oil, resin, silicon and salts (calcium oxalate or calcium carbonate).
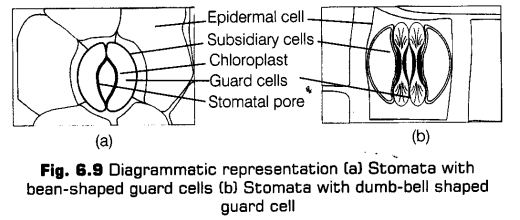
Stomata The stomata (sing, stoma) are openings in the epidermis of most of the aerial parts of the plants, especially the leaves. Each stomata is composed of two bean-shaped cells called as guard cells, which enclose stomatal pore. The guard cells are generally much smaller in size as compared to other epidermal cells. They are sensitive to even a small change in turgor pressure. The dimension of stomatal pore varies from species to species but it measures about 20 Jim long and about 10-20 p.m wide when fully open.
In some species, the guard cells are surrounded by subsidiary cells or accessory cells which differ morphologically from the other epidermal cells. The guard cell walls have special elastic properties. The adjoining cell walls of two guard cells around pore are free and not attached with each other.
These properties help them to stretch laterally during stomatal opening. The stomatal aperture, guard cells and the surrounding subsidiary cells are together called stomatal apparatus.
In most monocots, the guard cells are dumb bell-shaped. The stomata are mostly found on the upper epidermis of the leaves. In some hydrophytes, the stomata occur on the upper surface to avoid water contact.
Based on their distribution, stomata are of following types
(a) Apple Type Present on the under side of a leaf, e.g., Apple, mulberry.
(b) Oat Type Stomata are almost equal on the two surfaces, e.g, Maize, oat.
(c) Potato Type These are more on the under surface, e.g, Cabbage, potato, bean.
(d) Water lily Type These stomata are more on the upper surface, e.g., Many aquatic plants.
(e) Potamogeton Type Stomata vestigial or absent, e.g, Potamogeton.
Epidermal Appendages (Outgrowths)
The epidermis of most plants often bear outgrowth known as epidermal appendages or epidermal outgrowths. They are of following two types
(a) Trichomes The epidermal hairs present on the stem are called trichomes. These are epidermal outgrowths present temporarily or permanently on almost all plant parts. The trichomes can be further divided as hair, scales, colleters and water vesicles or bladders.
(b) Emergences (Prickles) They are multicellular, stiff and sharp epidermal outgrowths containing some inner tissues.
•They protect the pliant against excessive loss of water and grazing. They also helps in climbing in some plants, e.g., Rose.
Epidermal tissue system serves the following important functions
(a) It provides a protective covering all over the plant parts.
(b) It helps in gas exchange through stomata and lenticles present on the surface.
(c) The presence of cuticle helps in the reduction of evaporation of water (epidermis).
(d) The glandular trichomes excrete various useful plant products for the plant function.
(e) In some monocot leaves, the bulliform cells help in the rolling and unrolling of leaves. This property helps to reduce transpiration in xerophytic plants.
2. Ground Tissue System
All tissues, except epidermis and vascular bundles constitute the ground tissue system. It mainly forms the bulk of the plant body. It’s various components are hypodermis, cortex, endodermis, pericycle, medullary rays and pith.
i.Hypodermis This is the region situated just below the epidermis and as an outer region of cortex. It contains of one, two or few continuous or discontinuous layers of collenchyma (in dicots) or sclerenchyma (in monocots). It is protective and mechanical in function.
ii.Cortex The cortex lies between epidermis and endodermis consisting of parenchyma, collenchyma and sclerenchyma. The cortex is distinct in dicotyledons but not in monocotyledons. The cells of cortex contain starch grains, oil, tannins and crystals. Sometimes, cortical cells may contain chloroplasts and are called chlorenchyma.
In hydrophytes, the cortex may be aerenchymatous (Spongy tissue with large air spaces found between the cells of the stems and leaves of aquative plants). The special types of cells like sclereids, resin ducts, oil glands laticifers are found in this region. The cortex helps in performing vital functions, such as storage, etc.
iii. Endodermis This is the innermost layer of the cortex. It is single-layered, barrel-shaped and arranged without intercellular spaces. The cells are parenchymatous. The presence of bands of suberin on the radial and transverse wall is the characteristic feature. These bands are called casparian strips. The endodermal cells of roots usually have thick, radial and inner tangential walls.
These thick-walled cells form a continuous ring which is interrupted at certain places by passage cells, which are thin-walled and usually present opposite to the protoxylem region.
A well-developed endodermis is present in all types of roots, aerial stems of woody dicotyledons and gymnosperms with characteristic casparian thickenings.
The endodermis helps to control the movement of water and air between the cortex and xylem. It also helps to maintain the root pressure and conducts water to the protoxylem.
iv. Pericycle It is made up of a single layer or many layers of cells present between endodermis and vascular tissue. In roots, pericycle comprises cells of parenchyma. The pericycle is absent in roots and stems of some aquatic plants.
v. Medullary Rays The-medullary rays are non-vascular areas which occur between vascular bundles in dicot stems for lateral conduction. These are made up of parenchyma cells. These originate from the apical meristem. They serve the function of lateral transport.
vi. Pith The central portion of root and stem is occupied by pith. It contains parenchymatous cells and also sclerenchymatous cells laticifers, medullary vascular bundles, in some cases. In leaves, the ground tissue is parenchymatous and possesses chloroplast.
It performs the function of photosynthesis. The main function of pith is storage of water and food materials.
vii. Ground Tissue of Leaves In leaves the ground tissue of petiole is made up of parenchymatous cells with distinct intercellular spaces. In the lamina, the bulk of ground tissue is called mesophyll, which is usually differentiated into palisade and spongy parenchyma.
These cells are thin-walled and possess chloroplasts. The main function of mesophyll is in photosynthesis.
3. Vascular Tissue System
A vascular bundle is a strand of conducting tissue, which is generally composed of xylem and phloem in monocots and xylem, phloem and cambium in dicots.
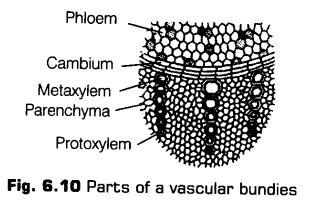
These tissues originate from the procambium and apical meristems. The arrangement of xylem and phloem is the characteristic to particular plant organs. However, a few exception are also there.
On the basis of arrangement of xylem and phloem in the vascular bundles, there are three types of bundles, i.e., radial, conjoint and concentric.
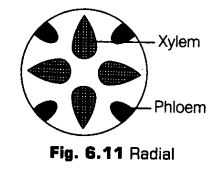
i.Radial The xylem and phloem alternate with each other separated by parenchymatous cells. This types of vascular bundles are called radial and is found mainly in roots.
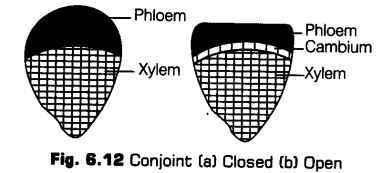
ii. Conjoint The xylem and phloem are present together in the same bundle on the same radius. Conjoint bundles are of two types, i.e., collateral and bicollateral.
(a) Collateral The xylem and phloem lie together on the same radius. The xylem lies inwards and the phloem outwards.
They are of two types
• In a dicot stem, the cambium is found to be present in between the xylem and phloem, such bundles are called open, e.g., Helianthus (sunflower).
* When the cambium is absent, the vascular bundle is called as a closed bundle, e.g., Zea mays (maize).
(b) Bicollateral This is the conjoint vascular bundle with two groups or patches of phloem, one on each side of the centrally located xylem. The various components are arranged in sequence of outer phloem, outer cambium, xylem, inner cambium and inner phloem. Such bundles are commonly found in the members of Cucurbitaceae. Such bundles are always open.
iii. Concentric A vascular bundle in which one tissue is completely surrounded by the other is called concentric. The concentric bundles are of two types, i.e., amphibasal (phloem lies in the centre and remains completely surrounded by xylem) and amphicribal (xylem lies in the centre and remains completely surrounded by phloem).
2 Anatomy of Dicotyledonous and Monocotyledonous Plants
The tissue organisation of roots, stems and leaves can be studied better and conveniently by the transverse sections of the mature zones of these organs.
I.Dicotyledonous Root
The primary internal structure of dicot root can be studied from the Transverse Section (TS) of a young root of sunflower, pea or gram. The primary root is the one which has only primary permanent tissues that are formed from vegetative shoot apex. Secondary tissues are absent.
The following structures can be seen from periphery towards the centre
1. Epiblema
It forms the outermost layer in young root. It is equivalent to epidermis of stem. The stomata and cuticle are not present in it. The cells are thin-walled and tubular. Some of the epiblema cells are prolonged to form thin-walled tubular structures called root hairs.
The cells which produce root hair are called root hair cells or trichoblasts. Due to the presence of root hairs, epiblema is also called piliferous layer (Pilus – hair; ferre – to carry) and rhizodertnis (Rhiza – root; derma – skin).
Root hairs having pectose layer on the outside, this is to help them to pass into the soil spaces for absorption of water and mineral salts. The active life span of root hairs is up to 7 days and die off in older parts of the root. The cell of older epiblema shrivel afterwards and become cutinised and suberised.
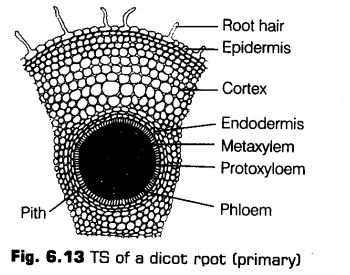
2. Cortex
It lies beneath the epiblema. It consists of several layers of thin-walled parenchymatous cells with conspicuous intercellular spaces. The cells of cortex store fipod. It also conducts water from the ebiblema to the inner tissues.
3. Endodermis
The innermost layer of the cortex is endodermis. It comprises of a single layer of barrel-shaped cells without any intercellular spaces. The endodermal cells are living and are rich in starch grains.
They have characteristic bands of thickenings along their radial and tangential walls. These are called casparian bands or casparian strips.
The casparian strips are made up of suberin and lignin. These strips prevent plasmolysis of endodermal cells and do not allow wall to wall movement of substances, between cortex and pericycle.
The cells of endodermis lying opposite to the protoxylem are thin-walled to permit free passage of water and minerals from cortex into the xylem. These are called passage cells.
4. Stele
All tissues on the innerside of the endodermis such as pericycle, vascular strand and pith constitute the stele.
i. Pericycle The next to endodermis lies a layer of thick-walled parenchymatous cells referred to as pericycle. The initiation of lateral roots and vascular cambium during the secondary growth takes place in these cells.
ii. Vascular Strand The vascular strand consists of separate bundles of xylem and phloem arranged alternately inner to the pericycle. Hence, the xylem and phloem bundles are equal in number and lie on different radii. Such vascular bundles are called radial bundles.
On the basis of number of xylem bundles, the root may be diarch (with two xylem bundles), triarch, tetarch, pentarch and polyarch (with more than five xylem bundles).
When the protoxylem is towards the periphery and the later formed xylem (metaxylem) is towards the centre of the root. This kind of xylem condition is called exarch and is characteristic of root.
The phloem and xylem bundles are separated from each other by one or more layers of small thin-walled cells called conjuctive tissue.
Later, it becomes meristematic and forms vascular cambium. The phloem tissues conducts organic food from leaf to the other parts of the plant. Secondary, growth occurs in dicot roots.
iii. Pith It is generally absent in dicot roots. If present, it is small. It consists of parenchyma cells that store food and waste products.
Features for Identification of Dicotyledonous Root
Dicotyledonous root can be easily identified with the followingfeatures
(i) Presence of root hairs.
(ii) Endodermis with casparian strips.
(iii) Absence of pith.
(iv) Radial bundles less than eight.
(v) Presence of exarch xylem.
II.Ionocotyledonous Root
This can be inferred from the following structures given below
1 Epiblema
It is the outermost, thin-walled, compactly arranged layer of cells. Some of the cells give rise to root hair. The root hair are unicellular and lie in contact with soil water. Both epiblema and root hair are devoid of cuticle. These helps in absorption of water and minerals. In older parts the epiblema is shed or impervious.
2 Cortex
It is a broad zone of parenchyma cells. The cells are thin-walled and enclose intercellular spaces. They normally store food. The cortex provides for radial movement of water and minerals from epiblema to the root interior.
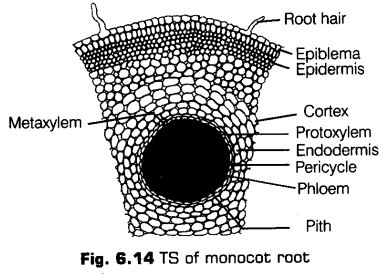
3 Endodermis
It is single-layered and made up of barrel-shaped cells which do not enclose intercellular spaces. The young endodermal cells possess an internal strip of suberin and lignin, which is known as casparian strip. Endodermal cells lying opposite the protoxylem groups however, remain in the primary stage with usual casparian strip.
These unthickened cells are called passage or transfusion cells. These cells helps in conduction of fluids and minerals from cortex into the xylem.
4. Stele
All tissues inside the endodermis, i.e., pericycle, vascular bundles and pith form the stele.
i. Pericycle It forms the outer boundary of stele. Pericycle may be uniseriate (single layered) or multiseriate (multilayered).
The pericycle does not form cambium. It only produces lateral roots. The pericycle is composed of thin-walled parenchymatous cells in a young root. Later, it becomes thick-walled in many monocot roots.
ii. Vascular strand Vascular strand is in the form of several alternate and radial xylem and phloem bundles. The vascular bundles are arranged in the form of a ring around a central pith.
The xylem bundles are exarch, i.e., protoxylem lies towards the outside while, the metaxylem faces inwards. Due to the pressure of numerous xylem bundles and exarch condition, the xylem of monocot root is polyarch.
Protoxylem vessels are narrow while, the metaxylem vessels are the broad. Xylem provides mechanical strength and helps in conduction of water and mineral salts.
Phloem and xylem are separated from each other by means of a narrow strip of conjuctive tissue. The phloem cells stored food, if parenchymatous. They provide mechanical strength on becoming sclerified. They are involved in the formation of cambium.
iii. Pith It is large and well-developed. It is large and made up of parenchymatous cells with intercellular spaces. These cells contain starch.
Features for Identification of Monocotyledonous Root
Monocotyledonous root can be easily identified with the following features
(i) Presence of root hairs.
(ii) Endodermis with passage cells.
(iii) Presence of pith.
(iv) Radial bundles more than eight.
(v) Xylem exarch.
(vi) Presence of an exodermis.
Differences between Dicot Root and Monocot Root

ysboiviedonous Stem
The transverse section (TS ) of a typical young dicotyledonous stem shows the following areas
1.Epidermis
The outermost protective layer of the stem is called epidermis. It is covered with a thin-layer of cuticle and may bear trichomes and a few stomata. The cuticle protects the tissues from injury as well as diseases from the entry of fungal spores and bacteria. It also helps to prevent loss of water.
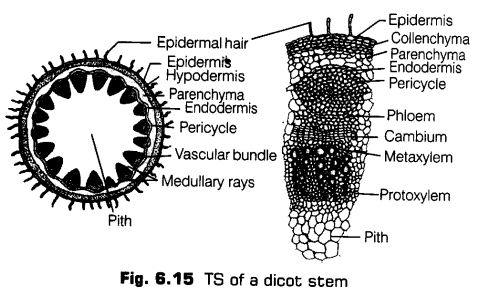
2.Cortex
This layer lies just below the epidermis and extends till endodermis. Its various parts are hypodermis, general cortex and endodermis.
Hypodermis It is just below the epidermis consisting of collenchymatous cells. The cells contain chloroplasts. It provides mechanical strength to the stem.
General Cortex It is located just below the hypodermis and consists of a few layers of parenchymatous cells. These cells are thin-walled and may contain chloroplasts.
Endodermis It lies just beneath the general cortex in the form of single layer of barrel-shaped cells surrounding the stele. It is the innermost layer of cortex. In sunflower, it contains starch, v hence is called starch sheath.
3. Pericycle
It exists between the endodermis and the vascular bundles. The cells are sclerenchymatous with lignified cell walls and a few parenchymatous cells dispersed in between. Each patch is associated with phloem of the vascular bundle and is called the hard bast.
4. Vascular Strand
The vascular strand consists of many vascular bundles, arranged in the form of a ring around a central pith and inner to pericycle.
Each vascular bundle consists of phloem (on the outside), xylem (towards the Inner side) with a strip of cambium, between the two.
The vascular bundles are thus, conjoint (i.e., consists of both xylem and phloem), collateral (i.e., phloem and xylem are on the same radius) and open (i.e., a strip of cambium present between the two).
5. Medullary or Pith Rays
These are non-vascular areas present in between the vascular bundles. The medullary rays connect pith with pericycle and cortex. Cells are larger than those of cortex. The medullary rays take part in radial conduction of materials, i.e., food, water, gases, etc.
6. Pith or Medulla
It consists of the central part of the stem. It consists of rounded, oval or polygonal parenchymatous cells. Intercellular spaces are absent. The cells store food materials and waste products.
Features for Identification of Dicotyledonous Stem
Dicotyledonous stem can be easily identified with the following features
(i) Occurrence of multicellular hair over epidermis.
(ii) Collenchymatous hypodermis.
(iii) Presence of bundle caps or sclerenchymatous pericycle over vascular bundles.
(iv) Endarch xylem.
lyionocqtyledonous Stem
The monocot stem possesses only primary structure. The different monocot stem from outside towards inside are consists of epidermis, hypodermis, ground tissue and vascular system.
1. Epidermis
It is single layered, having stomata in it. The cells have a thick cuticle layer on the outside.
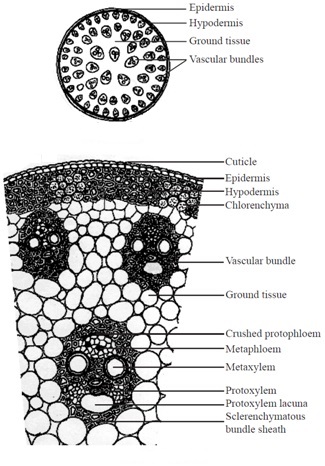
2. Hypodermis
It is 2-3 layered having lignified. sclerenchymatous cells present just below the epidermis.
3. Ground Tissue
It fills the whole interior of the stem containing parenchymatous cells. A number of vascular bundles are scattered in it.
4. Vascular System
Each vascular bundle in vascular strand is surrounded by a sheath of sclerenchyma known as bundle sheath cells. The vascular bundles possesses both phloem and xylem so, these are conjoint type.
The bundles are endarch with the protoxylem and metaxylem are arranged in the form. The divergent ends are occupied by two pitted vessels and convergent end by two smaller spiral vessels lying radially in the centre. A water containing cavity called lysigenous cavity is present in association with the protoxylem.
It is formed by the breakdown of inner protoxylem vessels and parenchyma during the earlier stages of growth. The cavity is absent or reduced in the smaller vascular bundles that occur in contact with sclerenchymatous hypodermis.
Features for Identification of Monocotyledonous Stem
Monocotyledonous stem can he easily identified with the following features
(i) Sclerenchymatous hypodermis present.
(ii) Undifferentiated ground tissue.
(iii) Vascular bundles scattered throughout ground tissue.
(iv) Vascular bundles are conjoint, collateral and closed.
(v) Protoxylem cavity present.
Dicotyledonous (Dorsiventral) Leaf
The dorsiventral leaves are generally horizontal and sunlight falls on their upper surface (ventral surface or adaxial surface).
The vertical section of a dorsiventral leaf through the lamina shows the following main parts
1. Epidermis
The epidermis covers both the upper (adaxial) and the lower (abaxial) surfaces of the leaf.
Upper Epidermis It is the uppermost, single layered, made up of parenchymatous cell, but sometimes, multilayered, e.g, Ficus, Piper, Nerium, Begonia. Also there is cuticle which covers the upper epidermis.
The outgrowths called papillae {e.g., Gladiolus) are sometimes present in epidermal cells. The stomata are usually less present in the upper surface. Chloroplasts are not present in this layer.
ii. Lower Epidermis The stomata and chloroplasts are more in number in the lower epidermis. There is sub-stomatal cavities present below the stomata for the gaseous exchange.
2. Mesophyll
It is differentiated in two parts in dorsiventral leaves, i.e., upper palisade and lower spongy parenchyma. The palisade cells contain abundant chloroplasts, Hence, they are the major seat of photosynthetic activity.
The spongy parenchyma lies below the palisade parenchyma and above the lower epidermis. This spongy parenchyma cells contain several chloroplasts but less than the number present in palisade cells.
Vascular System
The vascular bundles are conjoint, collateral, endarch and closed. Each bundle is surrounded by a bundle sheath of parenchymatous cells. The xylem is present towards upper epidermis (adaxial surface) and phloem towards lower epidermis (abaxial surface).
The xylem consists of vessels or trachae, tracheids, xylem parenchyma and xylem fibres. It is meant for the conduction of water and minerals.
The phloem is made up of sieve tubes, companion cells, phloem parenchyma and phloem fibres. Each vascular bundle is surrounded by a layer of thick-walled cells arranged compactly and known as bundle sheath cell (in C4-plants only).
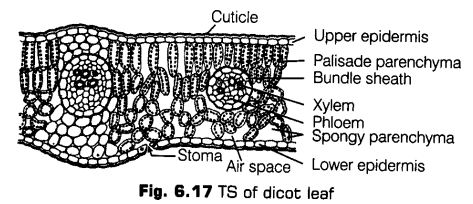
The vascular bundles can be seen in the veins and the midrib. The size of vascular bundles vary according to the size of the veins.
The veins vary in thickness in the reticulate venation.
Mesophyll is absent in the region of midrib and other larger veins. Collenchyma or sclerenchyma occur towards the two epidermal layers for providing mechanical strength. The centre contains a number of vascular bundles, which are embedded in a parenchymatous ground tissue.
Features for Identification of Dicotyledonous Leaf
Dicotyledonous leaf can be easily identified with the following features
(i) Bifacial flattered with stomata mostly on upper surface.
(ii) Mesophyll differentiated into palisade and spongy parenchyma. .
(iii) Vascular bundles with colourless bundle sheath (in C4-plants).
(iv) Vascular bundle with xylem towards upper side and phloem towards lower side.
The vascular bundles can be seen in the veins and the midrib. The size of vascular bundles vary according to the size of the veins.
The veins vary in thickness in the reticulate venation.
Mesophyll is absent in the region of midrib and other larger veins. Collenchyma or sclerenchyma occur towards the two epidermal layers for providing mechanical strength. The centre contains a number of vascular bundles, which are embedded in a parenchymatous ground tissue.
Features for Identification of Dicotyledonous Leaf
Dicotyledonous leaf can be easily identified with the following features
(i) Bifacial flattered with stomata mostly on upper surface.
(ii) Mesophyll differentiated into palisade and spongy parenchyma. .
(iii) Vascular bundles with colourless bundle sheath (in C4-plants).
(iv) Vascular bundle with xylem towards upper side and phloem towards lower side.
3.Vascular Bundle
A large number of vascular bundles are present, some of them are small and some are big. Each vascular bundle is surrounded by a bundle sheath of parenchymatous cells. Above and below the larger bundle, the patches of sclerenchymatous cells are present.
The vascular bundles are conjoint, collateral, endarch and closed. In some grasses, these are surrounded by a distinct parenchymatous bundle sheath. The xylem is present towards the upper epidermis and phloem towards the lower epidermis. The xylem and phloem elements of monocot leaves are similar to those of dicot leaves.
4. Midrib
It is the widest part of monocot leaf. A shallow groove is present in the upper or adaxial surface, while a broad ridge is present on the abaxial surface.
Features for Identification of Monocotyledonous Leaf
Monocotyledonous leaf can be easily identified with the following features
(i) Presence of large sized bulliform cells on upper surface.
(ii) Undifferentiated mesophyll.
(iii) Presence of bundle sheath with chloroplasts.
(iv) Vascular bundle with xylem towards upper side and phloem towards lower side.
(v) Xylem vessels rounded.
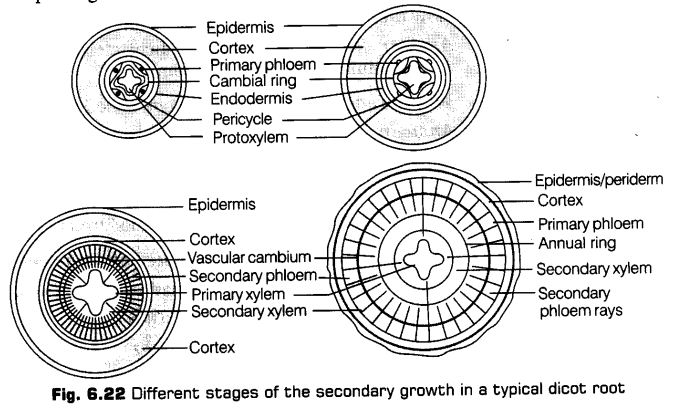
Secondary Growth
The growth of the roots and stems in length with the help of apical meristem is called the primary growth. Apart from primary growth, most dicot plants exhibit the increase in girth. This increase is called secondary growth.
Secondary Growth in Dicot Stem
In a dicot plant, secondary growth in stem occurs both in the stele and in the cortex. The dicot stem, in its primary state of growth contains narrow layers of intrafascicular cambium in between the xylem and phloem.
The tissues involved in the secondary growth are the two lateral meristems, i.e., vascular cambium and cork cambium.
1. Vascular Cambium
The meristematic layer that is responsible for cutting off vascular tissues such as xylem and phloem is called vascular cambium. It is present in a patch of a single layer in young stem which later on develops into a complete ring.
Formation of Cambium Ring
The parenchyma cells of the primary medullary rays adjacent to the intrafascicular cambium undergo dedifferentiation and give rise to interfascicular cambium. This joins the intrafascicular cambium of either side to form a complete ring of meristem called the cambium ring.
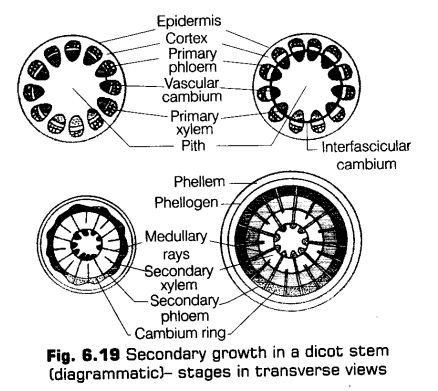
Activity of Cambial Ring
The cambial ring becomes active and begins to form new cells, both towards and inner and the outer sides. The cambial ring is made up of two types of cells ray initials and fusiform initials. The cells added to the inner side of cambium ring by the division of the fusiform initials gradually become the elements of the secondary xylem. While, the cells added to the outer side of the cambium become elements of the secondary phloem. While, the cells added by the division of ray initials to the inside as well as outside become elements of the secondary medullary rays.
The cambium is generally more active on the inner side than the outer. As a result, the amount of secondary xylem produced is more than secondary phloem and soon forms a compact mass.
The primary and secondary phloems get gradually crushed due to the continued formation and accumulation of secondary xylem. The primary xylem however, remains more or less intact, in or around the centre. At some places, the cambium forms a narrow band of parenchyma, which passes through the secondary xylem and the secondary phloem in the radial direction. These are secondary medullary rays.
Formation of Annual Rings
In tropical areas, the growth of secondary xylem is continuous. In others, yearly growth is quite distinct and appears in the form of annual rings. The transition from spring wood to autumn wood is gradual. After autumn wood and before spring wood of next year, there is no growth.
Therefore, change over from autumn wood to spring wood is sudden. The light coloured spring wood and its next dark coloured autumn wood constitutes an annual ring or growth ring.
It represents the total secondary xylem or wood formed in one year. Hence, by counting the number of annual rings, the age of a plant can be determined. This is done with the help of an instrument called increment borer. Besides giving the age of the plant, the annual rings can also provide information of the climatic conditions prevailing in the past.
Spring Wood and Autumn Wood
The activity of cambium is under the control of many physiological and environmental factors. In temperate regions, the climatic conditions are variable through the year.
In springs, cambium is very active and produces a large number of xylary elements having vessels with wider cavities. The wood formed in this season is called spring wood or early wood.
In autumn, the cambium is less active and forms few xylary elements that have narrow vessles. Thus, the wood formed is called autumn wood or late wood.
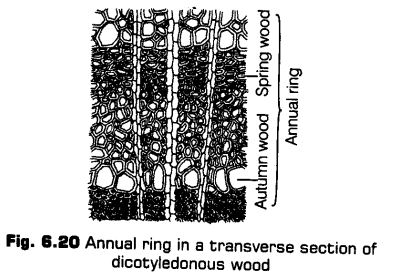
In old trees, the considerable region of secondary xylem is dark brown due to the accumulation of organic materials like tannins, resins, nils, gums, aromatic substances and essential oils in the central or innermost layers of the stem.
These substances make it hard, durable and resistant to the attacks of microorganisms and insects. This region comprises dead elements with highly lignified walls and is called heartwood.
The heartwood does not conduct water, but it gives mechanical support to the stem. The peripheral region of the secondary xylem, is lighter in colour and is known as the sapwood, which is involved in the conduction of water and minerals from root to leaf.
Cork Cambium
The stem continues to increase in girth due to the activity of vascular cambium. Due to this, the outer cortical and epidermis layers get broken and needs to be replaced to provide new protective cell layers. Therefore, another meristematic tissue called cork cambium or phellogen develops usually in the cortex region.
Phellogen is a couple of layers thick. It is made of narrow, thin-walled and nearly rectangular cells. Phellogen cuts off cells on both sides. The outer cells differentiate into cork or phellem while the inner cells differentiate into secondary cortex or phelloderm.
The cork is impermeable to water due to suberin deposition in the cell wall. The cells of secondary cortex are parenchymatous. The phellogen, phellem and phelloderm are collectively known as periderm.
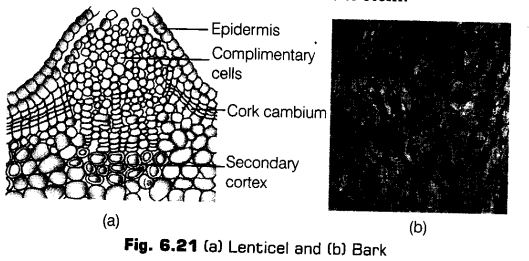
Bark
Bark is a non-technical term used to describe all tissues exterior to the vascular cambium, therefore including secondary phloem. The bark refers to a number of tissues, i.e., periderm and secondary phloem.
The bark that is formed early in the season is called early or soft bark. Towards the end of the season, late or hard bark is formed.
Lenticels
At certain regions of stem, the phellogen cuts off closely arranged parenchymatous cells on the outer side instead of cork cells. These parenchymatous cells soon rupture the epidermis, forming a lens-shaped openings called lenticels. The lenticel are mosdy found in woody trees.
The lenticels permit the exchange of gases between the outer atmosphere and the internal tissue of the stem.
Secondary Growth in Roots
The secondary growth in the root is the thickness due to the formation of secondary tissues by lateral meristems. With the exception of some annuals, most of the dicots and gymnosperms show secondary growth in their roots. It occurs by the production of two types of secondary tissues,
i.e., the secondary vascular tissues and periderm. These tissues are formed by meristems are vascular cambium and cork cambium, respectively.
Formation of Vascular Cambium
The conjunctive parenchyma cells, on the lateral sides of the phloem bundles as well as pericycle cells lying outside the protoxylem end becomes brick-shaped and meristematic. These develop into a wavy band of vascular cambium. The vascular cambium of the root is a secondary meristem. It continues to form secondary xylem on the inner side and secondary phloem on the outer side.
Secondary phloem consists of sieve tubes, companion cells, phloem parenchyma and phloem fibres. The secondary xylem contains elements like vessels, xylem parenchyma and xylem fibres.
Activity of Vascular Cambium
The vascular cambium derived from the pericycle gives rise to only ray cells. The formation of these ray cells is slower, than the formation of secondary vascular tissues. Due to this, the depressed parts of vascular cambium move outwardly and ultimately the cambium becomes circular.
Effect of Growth of Secondary Tissue
The primary phloem gets crushed due to the growth of secondary vascular tissues. The older secondary phloem is also partially destroyed as the new phloem becomes functional.
The primary and secondary xylems persist. Primary xylem is distinguishable by its : exarch nature and central position. As compared to the primary xylem, the vessels of the secondary xylem are broader and thinner. Annual rings are not very sharp.
This is because the climate of the soil does not vary much during different seasons.
Formation of Cork Cambium
Leave a Reply